
Chapter 8: Energy from Nutrients
8.8 Metabolism of Molecules Other Than Glucose
Lisa Bartee and Jack Brook
You have learned about the catabolism of glucose, which provides energy to living cells. But living things consume more than just glucose for food. How does a turkey sandwich, which contains various carbohydrates, lipids, and protein, provide energy to your cells?
Basically, all of these molecules from food are converted into molecules that can enter the cellular respiration pathway somewhere. Some molecules enter at glycolysis, while others enter at the citric acid cycle. This means that all of the catabolic pathways for carbohydrates, proteins, and lipids eventually connect into glycolysis and the citric acid cycle pathways. Metabolic pathways should be thought of as porous—that is, substances enter from other pathways, and other substances leave for other pathways. These pathways are not closed systems. Many of the products in a particular pathway are reactants in other pathways.
Carbohydrates
So far, we have discussed the carbohydrate from which organisms derive the majority of their energy: glucose. Many carbohydrate molecules can be broken down into glucose or otherwise processed into glucose by the body. Glycogen, a polymer of glucose, is a short-term energy storage molecule in animals (Figure 8.81). When there is plenty of ATP present, the extra glucose is converted into glycogen for storage (a process called glycogenesis). Glycogen is made and stored in the liver and muscle. Glycogen will be taken out of storage and converted to glucose if blood sugar levels drop. The break down of glycogen to glucose is called glycogenolysis. The presence of glycogen in muscle cells as a source of glucose allows ATP to be produced for a longer time during exercise.
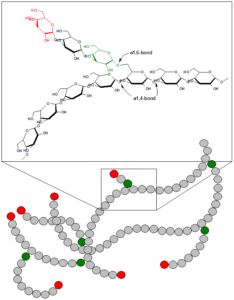
The amount of glycogen we can store in the body is limited. After a high-carbohydrate meal, our glycogen stores will reach capacity fairly quickly. After glycogen stores are filled, glucose will have to be metabolized in different ways for it to be stored in a different form, often as fat.
Most other carbohydrates enter the cellular respiration pathway during glycolysis. For example, sucrose is a disaccharide made from glucose and fructose bonded together. Sucrose is broken down in the small intestine. The glucose enters the beginning of glycolysis as previously discussed, while fructose can be slightly modified and enter glycolysis at the third step. Lactose, the disaccharide sugar found in milk, can be broken down by the lactase enzyme into two smaller sugars: galactose and glucose. Like fructose, galactose can be slightly modified to enter glycolysis.
Because these carbohydrates enter near the beginning of glycolysis, their catabolism (breakdown) produces the same number of ATP molecules as glucose.
Proteins
Proteins are broken down by a variety of enzymes in cells. Most of the time, amino acids are recycled into new proteins and not used as a source of energy. This is because it is more energy efficient to reuse amino acids rather than making new ones from scratch. The body will use protein as a source of energy if:
- There are excess amino acids (you consume a lot of protein)
- The body is in a state of famine (you are starving and have no other source of energy available)
When proteins are used in the cellular respiration pathway, they are first broken down into individual amino acids. The amino group from each amino acid is removed (deaminated) and is converted into ammonia. In mammals, the liver synthesizes urea from two ammonia molecules and a carbon dioxide molecule. Thus, urea is the principal waste product in mammals from the nitrogen originating in amino acids, and it leaves the body in urine.
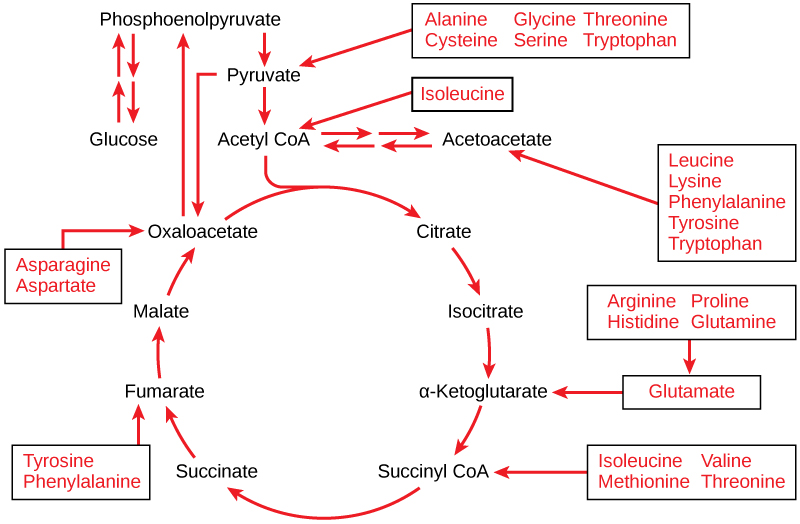
Once the amino acid has been deaminated, its chemical properties determine which intermediate of the cellular respiration pathway it will be converted into. These intermediates enter cellular respiration at various places in the Citric Acid Cycle (Figure 8.82).
Gluconeogenesis
Gluconeogenesis is the synthesis of glucose from non-carbohydrate sources. Gluconeogenesis is basically glycolysis in reverse, with minor changes. The glycerol from triglycerides can also be used for gluconeogenesis, although fatty acids cannot. Certain amino acids can be used for this process.
Not all amino acids can be used for gluconeogenesis. The ones that can be used are termed glucogenic, and can be converted to either pyruvate or a citric acid cycle intermediate. Other amino acids can only be converted to acetyl-CoA or acetoacetyl-CoA, which cannot be used for gluconeogenesis. However, acetyl-CoA or acetoacetyl-CoA can be used for ketogenesis to synthesize the ketone bodies, acetone and acetoacetate. Thus, these amino acids are instead termed ketogenic. Ketone bodies can be used to form fatty acids for energy storage. Some amino acids can be either ketogenic or glucogenic, depending on the body’s needs.
Lipids
Triglycerides (fats) are a form of long-term energy storage in animals. Triglycerides store about twice as much energy as carbohydrates. Triglycerides are made of glycerol and three fatty acids. Glycerol can enter glycolysis. Fatty acids are broken into two-carbon units that enter the citric acid cycle (Figure 8.83).
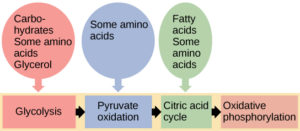
Remember that if oxygen is not available, glycolysis can occur but not the citric acid cycle or oxidative phosphorylation. Since fatty acids enter the pathway at the citric acid cycle, they can not be broken down in the absence of oxygen. This means that if cells are not performing aerobic cellular respiration, the body can not burn fat for energy. This is why posters about the “Fat Burning Zone” in a gym specify that you need to have a lower heart rate / breathing rate to burn more fat – cells that are not doing aerobic respiration can’t burn fat for fuel!
Lipogenesis & Lipolysis[1]
Lipogenesis is the synthesis of fatty acids, beginning with acetyl-CoA. Remember that acetyl-CoA is the product of the transition reaction that is the starting point of the citric acid cycle. Acetyl-CoA moves out of the mitochondria and is subsequently combined with additional acetyl-CoA molecules to form palmitate, a 16-carbon fatty acid.[2] The palmitate produced can be used as a component in the production of triglycerides (fat) for storage.
Lipolysis is the cleavage of triglycerides to glycerol and fatty acids. There are two primary lipolysis enzymes: lipoprotein lipase (LPL) and hormone-sensitive lipase (HSL). Despite performing the same function, the enzymes are primarily active for seemingly opposite reasons.
In the anabolic state (after eating a meal), LPL on the lining of blood vessels cleaves lipoprotein triglycerides into fatty acids so that they can be taken up into adipocytes (fat cells) for storage as triglycerides, or muscle cells where they are primarily used for energy production.
HSL is an important enzyme in adipose tissue (fat), which is a major storage site of triglycerides in the body. HSL activity is increased by glucagon and epinephrine (“fight or flight” hormone), and decreased by insulin. Thus, during hypoglycemia (such as during a fast; a catabolic state), or a “fight or flight” response, triglycerides in the adipocytes (fat cells) are cleaved, releasing fatty acids into circulation. The free fatty acids bind with a transport protein that carries them to muscle cells for use as an energy source. Thus, HSL is important for mobilizing stored fatty acids so they can be used to produce energy.
Catabolism refers to catabolic metabolic pathways. Catabolic pathways break down polymers into their monomers, releasing energy
Amino acids are the building blocks of proteins, simple subunits composed of carbon, oxygen, hydrogen, and nitrogen.
Glucogenic amino acids , and can be converted to either pyruvate or a citric acid cycle intermediate.
Ketogenic amino acids can only be converted to acetyl-CoA or acetoacetyl-CoA, which cannot be used for gluconeogenesis.
The electron transport chain and the production of ATP through chemiosmosis are collectively called oxidative phosphorylation.
Aerobic respiration refers to the process by which cells make adenosine triphosphate (ATP) in the presence of oxygen. This is the reason why we breathe oxygen in from the air. Aerobic respiration involves glycolysis, the citric acid cycle, and the electron transport chain / chemiosmosis.
Glucagon communicates to the cells in the body to stop using all the glucose. More specifically, it signals the liver to break down glycogen and release the stored glucose into the blood, so that glucose levels stay within the target range and all cells get the needed fuel to function properly.
A hormone is a compound that is produced in one tissue, released into circulation, then has an effect on a different organ.
Insulin is a hormone released from the pancreas that brings blood glucose levels down. Insulin sends a signal to the body’s cells to remove glucose from the blood by transporting it into different organ cells around the body and using it to make energy. In the case of muscle tissue and the liver, insulin sends the biological message to store glucose away as glycogen.
Hypoglycemia is when blood glucose levels are lower than normal. Hypoglycemia can lead to feelings of tiredness, anxiety, or irritability, and if not treated, can progress to seizures and lack of consciousness, as the brain is unable to function without glucose as an energy source.

{kind=link}